the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 04 Jul 2022
| 04 Jul 2022
A validation standard for area of habitat maps for terrestrial birds and mammals
Prabhat Raj Dahal
Maria Lumbierres
Stuart H. M. Butchart
Paul F. Donald
Carlo Rondinini
Area of habitat (AOH) is a deductive model which maps the distribution of suitable habitats at suitable altitudes for a species inside its broad geographical range. The AOH maps have been validated using presence-only data for small subsets of species for different taxonomic groups, but no standard validation method exists when absence data are not available. We develop a novel two-step validation protocol for AOH which includes first a model-based evaluation of model prevalence (i.e, the proportion of suitable habitat within a species' range), and second a validation using species point localities (presence-only) data. We applied the protocol to AOH maps of terrestrial birds and mammals. In the first step we built logistic regression models to predict expected model prevalence (the proportion of the range retained as AOH) as a function of each species' elevation range, mid-point of elevation range, number of habitats, realm and, for birds, seasonality. AOH maps with large differences between observed and predicted model prevalence were identified as outliers and used to identify a number of sources of systematic error which were then corrected when possible. For the corrected AOH, only 1.7 % of AOH maps for birds and 2.3 % of AOH maps for mammals were flagged as outliers in terms of the difference between their observed and predicted model prevalence. In the second step we calculated point prevalence, the proportion of point localities of a species falling in pixels coded as suitable in the AOH map. We used 48 336 141 point localities for 4889 bird species and 107 061 point localities for 420 mammals. Where point prevalence exceeded model prevalence, the AOH was a better reflection of species' distribution than random selection. We also found that 4689 out of 4889 (95.9 %) AOH maps for birds, and 399 out of 420 (95.0 %) AOH maps for mammals were better than random. Possible reasons for the poor performance of a small proportion of AOH maps are discussed.
- Article
(3108 KB) - Full-text XML
- BibTeX
- EndNote





An accurate estimate of the distribution of species is central to ecological and conservation research and action. There are three different classes of information on the distribution of species (Rondinini and Boitani, 2006). These are (1) point localities (latitude and longitude) of individuals, (2) geographic ranges, which are derived by mapping the extent of known point localities along with expert knowledge and (3) species distribution models, which use environmental and other relevant variables associated with the species to refine geographical ranges. Species distribution models are of two types (Stoms et al., 1992). The first are deductive models, which use expert-based information on species' habitat use to model the suitable areas for the species. The second type are inductive models, in which the environmental conditions at point localities where the species were recorded are interpolated over wider areas.
Area of habitat (AOH; also known as extent of suitable habitat, ESH) is a deductive model which maps the distribution of suitable habitats for a species inside its broad geographical range (Brooks et al., 2019). It aims to reduce commission errors present in the range map while minimizing omission errors. Several sets of AOH maps for different taxonomic groups at continental and global scales have already been produced (Rondinini et al., 2005; Rondinini and Boitani, 2006; Catullo et al., 2008; Jenkins and Giri, 2008; Rondinini et al., 2011; Ficetola et al., 2015; Tracewski et al., 2016; Lumbierres et al., 2021b).
Habitat models are prone to two major types of errors: omission errors occur when suitable habitat areas for the species are wrongly mapped as being unsuitable, commission errors occur when areas unsuitable for the species are wrongly mapped as being suitable. Quantification of these errors is one of the key parts of the habitat modeling process and is done by validation. The omission and commission errors could both be quantified only when independent presence and absence data on the species are available. In such cases standard validation metrics such as true skill statistics (TSS) (Allouche et al., 2006) and the Boyce Index (Boyce et al., 2002) are used. In the case of AOH maps produced for large taxonomic groups when true absence data are not available, no standard validation method exists.
Rondinini et al. (2011) and Ficetola et al. (2015) used point localities from GBIF (Global Biodiversity Information Facility) (https://www.gbif.org, last access: 1 May 2020) to validate AOH maps for mammals and amphibians. AOH maps for South Asian mammals (Catullo et al., 2008) and African vertebrates (Rondinini et al., 2005) were also validated using point localities. Brooks et al. (2019) recommend using point localities for validation and inclusion of AOH maps for IUCN (International Union for Conservation of Nature) Red List assessment. However, point localities are often not available for many species and are biased towards certain taxonomic group and well-studied areas.
In this paper, we developed a novel two-step validation protocol for AOH which includes: (a) a model-based evaluation of model prevalence (i.e., the proportion of a species' range that comprises AOH), and (b) a validation using species point localities (presence-only) data. We demonstrate the use of this approach by validating a new set of AOH maps produced by Lumbierres et al. (2021b) for all terrestrial birds and mammals. The validation method developed here is an iterative process whereby systematic errors in the production of AOH (e.g., in the matching of habitat classes to land cover maps) were identified using logistic regression models, then corrected where possible and a new set of AOH maps produced. Then we employed a point validation analysis for the subset of species for which point localities were available to assess the performance of the AOH maps. Finally, we assessed the extent to which the subset of species for which point locality data were available were representative of those for which no point data were available.
2.1 Identifying optimal threshold for the habitat-land cover model to produce AOH maps
The new set of AOH maps (Lumbierres et al., 2021b) was produced at a resolution of 100 m using a novel habitat-land cover model (Lumbierres et al., 2021a) which associated the different land cover classes in the Copernicus land cover map (Buchhorn et al., 2019) with the Level-1 habitat classes of the IUCN habitat classification scheme (IUCN, 2012). The IUCN habitat classification scheme is a hierarchy of habitat classes, and each species assessed in the IUCN Red List is assigned to one or more of these habitat classes, based on available information in the literature, unpublished reports and expert knowledge. The habitat-land cover model (Lumbierres et al., 2021a) has the provision of associating IUCN habitat classes to land cover classes using three different thresholds (1, 2 and 3). Lower thresholds permit weaker associations between land cover and habitat classes. Therefore, with threshold 1 each land cover class is associated with more habitat classes than with threshold 3. Lumbierres et al. (2021b) produced a set of AOH maps for each of the three different thresholds by clipping out of each species range any cells of land cover that were not linked by the model to the habitat class(es) to which the species was coded, then further clipping out parts of the range falling outside the elevation range of the species.
In order to identify the best threshold among the three thresholds and to validate the set of AOH maps with the best threshold at species level, we quantified two measures: “model prevalence” and “point prevalence”. Model prevalence is defined as the proportion of pixels inside the range that were retained in the AOH. For example, if 25 % of the pixels present in the original range map are clipped out because they contain unsuitable habitat, fall outside the species elevation range or both, the model prevalence is 0.75. Point prevalence is defined as the proportion of point localities (or their buffers) out of all points inside the range of a species falling inside the suitable pixels. For example, the red-tailed comet (Sappho sparganurus) had a total of 71 point localities within its range, of which 62 fell in pixels coded as suitable in the species AOH map, giving a point prevalence of = 0.88.
Because the number of habitats associated with each land cover class decreases with increasing thresholds, model prevalence is highest for threshold 1 models and lowest for threshold 3 models. With increasing thresholds, commission errors are expected to decrease (which is the main purpose of AOH) but omission errors might increase. Our validation protocol therefore aimed to control for omission errors. We did this by calculating point prevalence and model prevalence across the three thresholds and identified the set of AOH maps for which the mean model prevalence was lowest without compromising the mean point prevalence.
The point localities for bird species were downloaded from eBird (https://www.ebird.org, last access: 1 May 2020), the largest global repository for data on point localities of birds. eBird provides a metadata file called “eBird basic data set” (Cornell Lab of Ornithology, 2020) which is a compilation of all the validated point localities at species level and is updated monthly. These point localities are submitted by citizen scientists as well as experts worldwide and are checked by local experts to remove obvious misidentifications before they are made available for download (Sullivan et al., 2009). We first downloaded the metadata file from eBird updated in January 2020 which was then queried in R (R Core Team, 2018) using the auk package (Strimas-Mackey et al., 2018), as recommended by eBird, to extract the point localities at species level. The taxonomy of Birdlife International (BirdLife International and Handbook of the Birds of the World, 2020), which is that followed by the IUCN, was matched with eBird's taxonomy and point localities of only those species common to both were queried and extracted from the metadata. Of the 10 813 species listed in Birdlife International's list for which AOH maps were produced, 9628 species matched by name. Of these 9628 species, 8998 species shared the same taxonomic concept and for 730 species the scientific names matched but the taxonomic concept did not.
To ensure that only high-accuracy points were used for the validation, we selected the stationary points from eBird's metadata. The stationary points are those that have coordinate uncertainty of less than 30 m. We then applied a temporal filter of 2019–2020 because the point localities from 2005–2018 were used to calibrate the habitat-land cover model in Lumbierres et al. (2021a). This ensured there was no overlap between the calibration and validation data. The points were further filtered by the range polygon of the species provided by the IUCN Red List website (IUCN, 2020) to remove the small number of points falling outside the range (many of them likely to be misidentifications). Since the AOH maps in question only include a certain combination of presence, origin and seasonality of the range, we used the same combination to filter the point localities. This ensured that we only included points which fell inside the boundaries of the selected range maps. We also made sure that only one point locality was allowed per pixel of the AOH map to avoid clustering of points. Finally, we excluded species which had fewer than 10 point localities after all the filters were applied. A total of 4889 bird species had 4 836 141 point localities after filtering. For mammals, point localities were downloaded from GBIF (GBIF.org, 2021) following the taxonomy of Global Mammal Assessment (which is followed by IUCN) with the same temporal and spatial filters as with birds except the filter of coordinate uncertainty which was set to 300 m for mammals. This was done because far too many mammal species would be excluded in the validation if we only considered point localities with coordinate uncertainty of less than 30 m. The rgbif package (Chamberlain et al., 2021) in R was used to download the points for mammals. A total of 107 061 point localities for 420 species were available for mammals after applying all the filters.
A buffer of 300 m was applied around all the point localities to account for the positional uncertainty of the points and for the fact that the location usually records that of the observer at the time of observation and not the focal animal, following Jung et al. (2020). The buffers of point localities were then overlaid on top of the AOH maps across all three thresholds at species level and if at least one pixel coded to suitable habitat was found inside the buffer, the pixel was considered to be validated at that point locality. The count of validated pixels was used to calculate point prevalence at species level across all three thresholds.
We identified the threshold that produced a set of AOH maps for which the mean model prevalence was lowest without detriment to the mean point prevalence.
We then employed a two-step approach to validate the set of AOH maps with the optimal threshold. In the first step, we identified potential systematic errors in the AOH maps using a modeling approach that aimed to identify species whose model prevalence was larger or smaller than expected, given the characteristics of the species concerned. In the second step, we validated the AOH maps using point localities following Rondinini et al. (2011).
2.2 A modeling approach to identify outliers
We used logistic generalized linear models to predict model prevalence of the set of AOH maps produced using the optimal threshold as a function of a number of independent variables, and identified outliers whose observed model prevalence was significantly higher or lower than predicted by the model. Outliers were then examined to identify systematic errors in, for example, the way habitats were coded to land cover classes in the production of the AOH maps, and to identify species that might be coded to the wrong habitats or elevation limits. For example, if a species' range includes a high proportion of a particular land cover type not associated with the suitable habitats of the species in the habitat-land cover association table (Lumbierres et al., 2021a), or if errors in coding species to elevation limits mean that most of the range is outside the species' stated limits, the model prevalence would be lower than predicted by the model.
The predictors fitted to the logistic models included elevation range of the species (upper elevation limit minus lower elevation limit), mid-point of the elevation range, number of habitats to which the species is coded against in the IUCN Red List, seasonality of species (breeding and non-breeding ranges in case of migratory birds) and the geographical realm of the species. In the case of migratory birds, Lumbierres et al. (2021b) has three different classes (resident, breeding and non-breeding seasonalities) of AOH maps based on seasonality of the species. We merged resident seasonality to breeding and non-breeding seasonalities to have AOH maps with only two seasonalities (breeding and non-breeding). The dependent variable was the model prevalence of the AOH maps. Data from a total of 10 475 AOH maps for 9163 bird species (including for some species with separate breeding and non-breeding ranges) and 2758 AOH maps for 2758 mammal species were used to build logistic regression models for birds and mammals separately using the lme4 (Bates et al., 2015) package in R. Data on elevation were lacking for many mammal and bird species which is the reason why not all species could be included in the logistic model. After testing taxonomic genus, family and order as random effects in the model to control the non-independence of closely related taxa, family was selected for fitting as the residual variance was lowest for the models with family as the random effect for both birds and mammals. The predictive power of the model was assessed by calculating marginal R2 and conditional R2 using the insight (Lüdecke et al., 2019) package in R. The marginal R2 expresses how much of the variation in data is explained by the fixed effects and conditional R2 tells how much of the variation in data is explained by both fixed and random effects.
The Tukey fences outlier detection test (Wilcox, 2017) was used to identify outliers based on the difference between the estimated and observed values of model prevalence. This test uses the interquartile ranges to estimate the outliers in a data-set. The outlier test identified mild lower and upper threshold values for the difference between estimated and observed values.
The AOH maps identified as mild upper outliers have an observed model prevalence much larger than their predicted model prevalence, whereas maps identified as mild lower outliers have an observed model prevalence much smaller than their predicted model prevalence.
In order to investigate the sources of errors in the outliers, we produced two more sets of AOH maps for the outliers. One set included AOH maps which were produced by clipping the range of the species by the altitudinal range only (AOHElevation only). Similarly, the other set included AOH maps which were derived by clipping the range with only suitable habitat of the species (AOHHabitat only). If the model prevalence of an outlier was equal or nearly equal to the model prevalence of its AOHElevation only, then we concluded that the underrepresentation of model prevalence could be attributed to errors in the elevation range of the species. If the model prevalence of an outlier was equal or nearly equal to the model prevalence of AOHHabitat only, then the source of error could be attributed to the mapping of the habitats inside the range using the habitat-land cover crosswalk (Lumbierres et al., 2021a) or to errors in the species' habitat coding. Furthermore, in some of the outliers the underrepresentation could result from inclusion of a large proportion of habitats which were unsuitable for the species but were inside the range map of the species. Outliers do not necessarily represent errors in AOH, as species might legitimately have very high or low model prevalence, but by identifying suites of outliers sharing common characteristics we were able to identify and correct a number of systematic errors in AOH production. The models also allowed us to identify species for which AOH maps might be unreliable and the habitat and elevation coding needs to be checked.
2.3 Point validation of AOH maps of terrestrial birds and mammals
We validated 4889 bird and 420 mammal species' AOH maps using the filtered point localities. The point validation was done by comparing the model and point prevalence at species level. If the point prevalence exceeded model prevalence at species level, the AOH maps performed better than random, otherwise they were no better than random. We also calculated the percentage of suitable habitat pixels inside the buffers to ensure that the validation success was not due to one or few pixels falling inside the 300 m buffer.
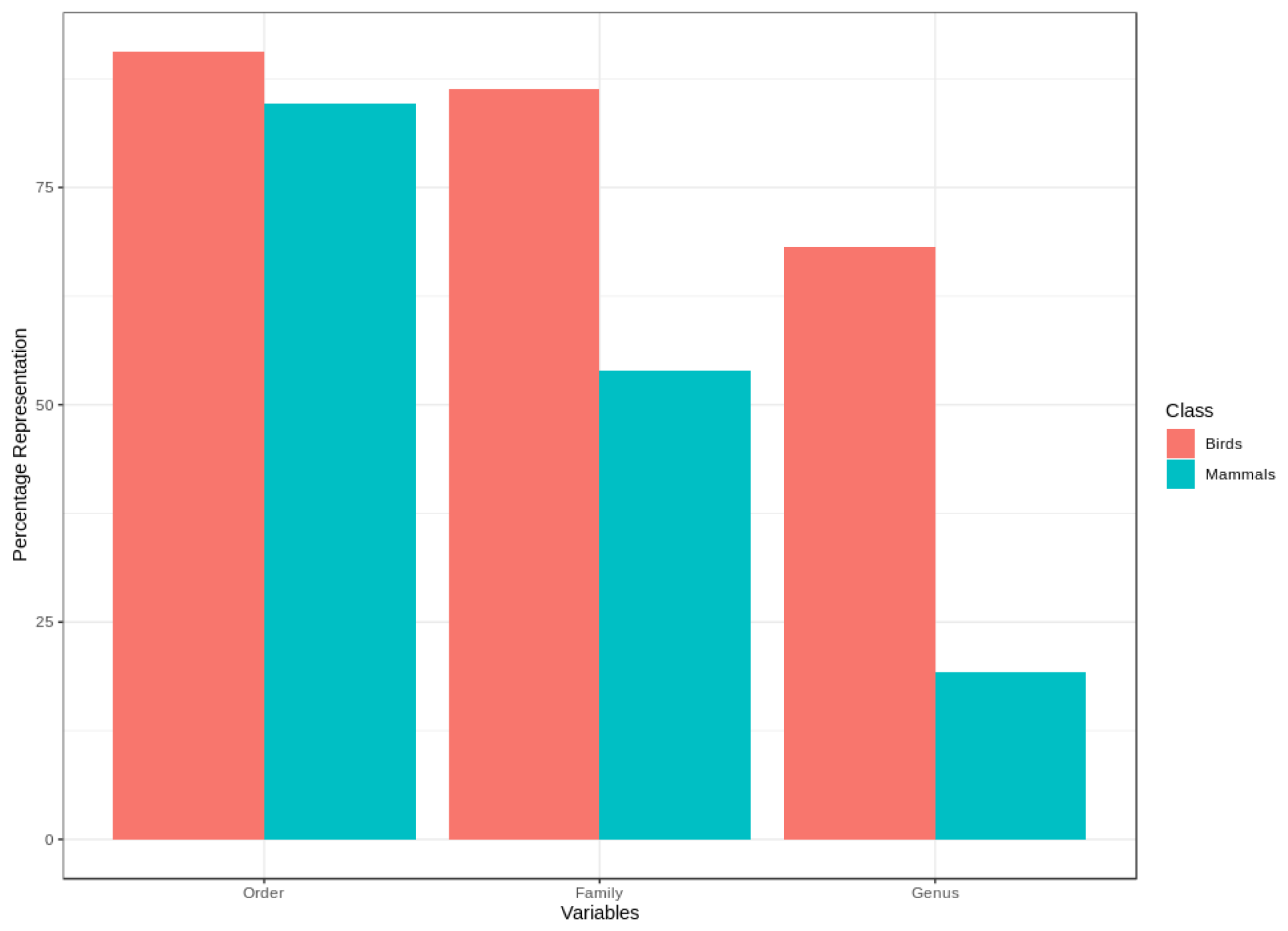
One of the major issues with citizen science data is that there is often a nonrepresentative spread of data across species. It is therefore possible that the species included in the point validation analysis are not representative of the species not included. We assessed how representative the validation sample size was by comparing the representation of variables, such as family, order, genus, realm, elevation range, mid-point of the elevation range, range size and extinction risk categories for birds and mammals between species with and without point data. The point validation was done in R and GRASS (GRASS Development Team, 2017).
After comparing point and model prevalence of 4889 birds and 420 mammal species across all 3 thresholds, we selected the set of AOH maps derived by using threshold 3 in the habitat-land cover model. At threshold 3, the mean model prevalence decreased as compared to thresholds 1 and 2 with much lower change in the mean point prevalence (Tables 1 and 2) for both birds and mammals.
Table 1Mean model and point prevalence for AOH maps with standard deviation of 4889 bird species across 3 different thresholds.
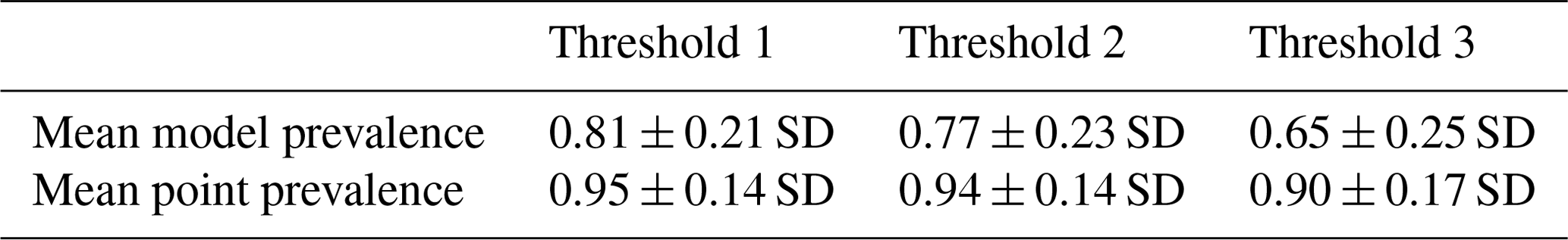
Table 2Mean model and point prevalence for AOH maps with standard deviation of 420 mammal species across 3 different thresholds.

We also assessed the relative contribution of elevation range, habitat, and both in reducing the range to AOH. For both birds and mammals, most of the pixels removed from the range were because either the habitat or the elevation was unsuitable, with a relatively small proportion being removed because both were unsuitable (Figs. 1, 2). The proportion of the range that was clipped out on the basis of having unsuitable habitat at suitable elevations increased as model prevalence decreased, whereas there was little change across the same axis in the proportion of the range that was excluded on the basis of having suitable habitat at unsuitable elevations (Figs. 1, 2). The number of both bird and mammal species peaked at model prevalence of 95 %–100 % and gradually decreased as the model prevalence decreased.
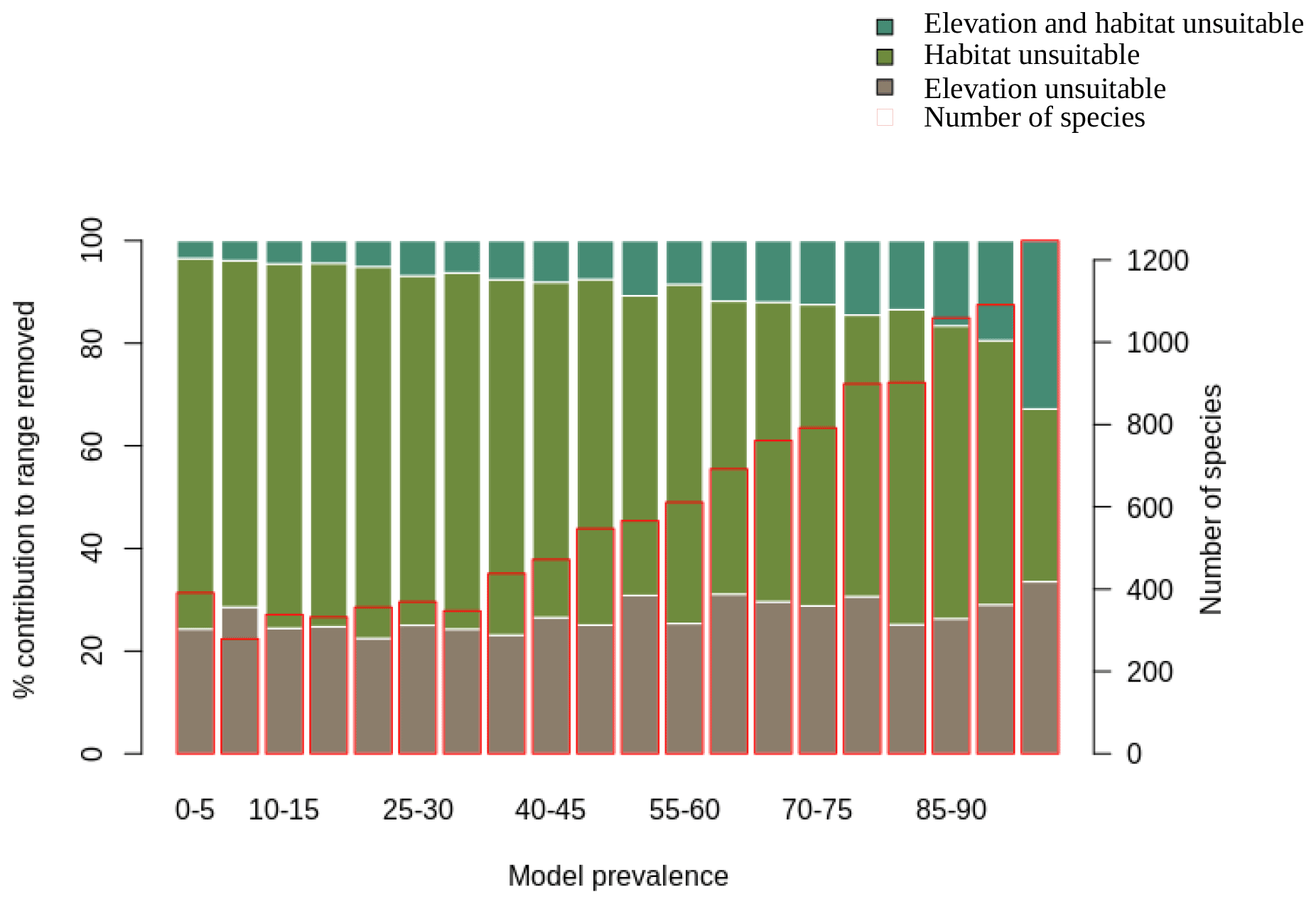
Figure 1Percentage contribution of elevation range, habitat and both in clipping the IUCN range to produce AOH maps for birds. Each bar represents a 5 % bin of model prevalence, divided to show how much of the range was clipped out due to unsuitable habitat at suitable elevations (“habitat unsuitable”), by suitable habitat at unsuitable elevations (“elevation unsuitable”) and by unsuitable habitat at unsuitable elevations (“elevation and habitat unsuitable”). The red blocks correspond to the second y-axis and show the number of species falling into each 5 % bin of model prevalence.
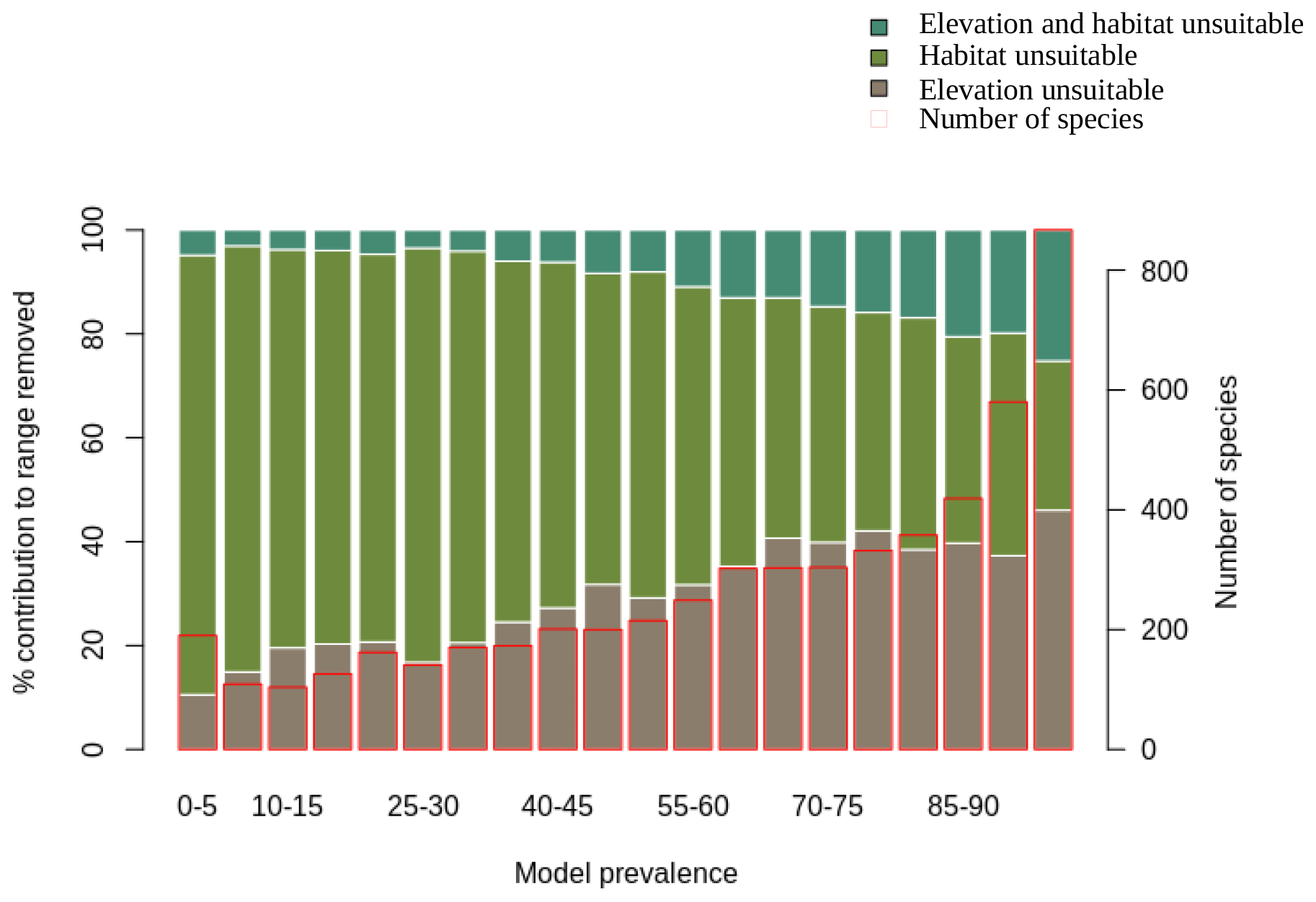
Figure 2Percentage contribution of elevation range, habitat and both in clipping the IUCN range to AOH for mammals. See caption to Fig. 1 for interpretation.
For birds, the logistic model identified 178 AOH maps (1.7 %) as lower outliers and 118 AOH maps (1.1 %) as upper outliers out of 10 475 AOH maps for 9163 terrestrial bird species. Similarly for mammals, the logistic model was applied to the AOH maps of 2758 species and identified 64 (2.3 %) as lower outliers and 21 (0.8 %) as upper outliers.
The mean of mid-point of elevation of the bird and mammal species identified as upper outliers was 2725 and 3193 m, respectively, while the mid-point of elevation for species which were not identified as upper outliers was 1261 m for birds and 1289 m for mammals. This suggests that species identified as upper outliers were those found in higher elevation. These species were identified as upper outliers because the logistic models predicted low model prevalence at higher elevations. Also, the range maps for high-altitude species are drawn using contour maps, therefore most of the range is within the correct altitudinal band leading to high model prevalence for these species.
The lower outliers indicate where model prevalence was possibly underestimated due to potential errors in habitat mapping and/or coding and elevation range of the species. We found that the habitats “shrubland” and “savannah” in the habitat-land cover crosswalk were not associated with the land cover class “herbaceous cover”, leading to underrepresentation of these habitat types and hence lower model prevalence than estimated by the logistic model (Fig. A1 in Appendix). We also found mismatch in the elevation range and geographical range for the lower outliers (Fig. A2). There were few cases where the range included a large proportion of a particular land cover type which was not associated with the suitable habitat of the species (Fig. A3). Moreover, we found that there was no land cover information in the Copernicus land cover map for very small range polygons located on oceanic islands which caused the AOH maps for these species to be empty. Furthermore, the land cover class “open forest unknown” was discarded in the habitat-land cover model. This led to low model prevalence of AOH maps for some species whose ranges included this land cover. This was corrected and a new set of AOH maps produced.
3.1 Point validation
Out of 4889 bird species (45 % of all bird species) for which point data were available, 4689 (95.9 %) had higher point prevalence than model prevalence and 200 species had a lower point prevalence than model prevalence (Fig. 3). The mean percentage of pixels coded as suitable inside the 300 m buffers of point localities of 4889 species of birds was 62 % (Fig. A5).

Figure 3Point prevalence vs. model prevalence for terrestrial birds. Colors indicate the number of habitats each species is coded to, size of circles indicates the number of point localities.
Out of 420 mammal species (8 % of all mammal species) for which point data were available, 399 (95.0 %) had a point prevalence higher than model prevalence (Fig. 4). The mean percentage of pixels coded as suitable inside the 300 m buffers of point localities of 420 species of mammals was 78 % (Fig. A5).
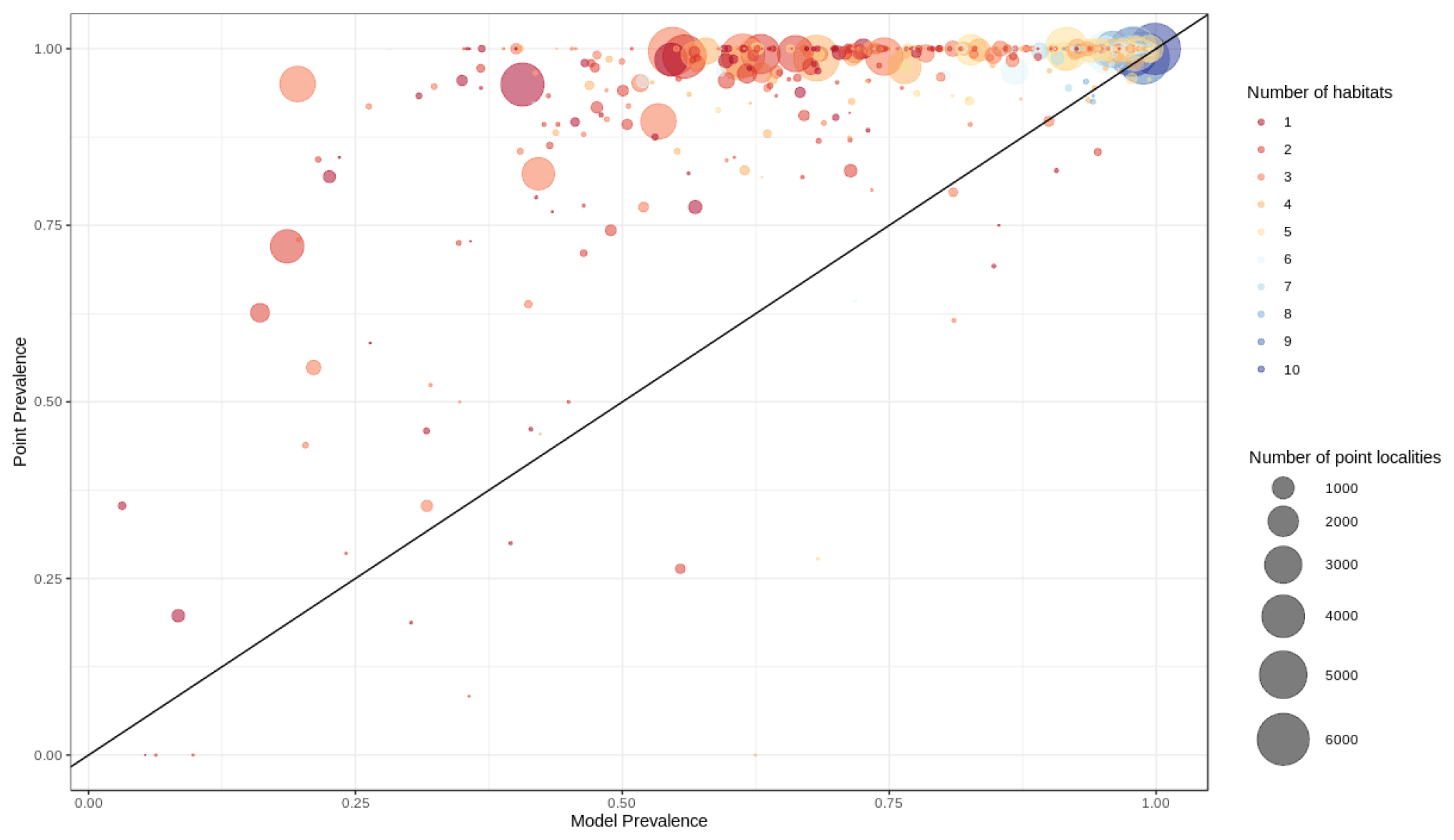
Figure 4Point prevalence vs. model prevalence for terrestrial mammals. Interpretation as in Fig. 3.
3.2 Representativeness of validation sample
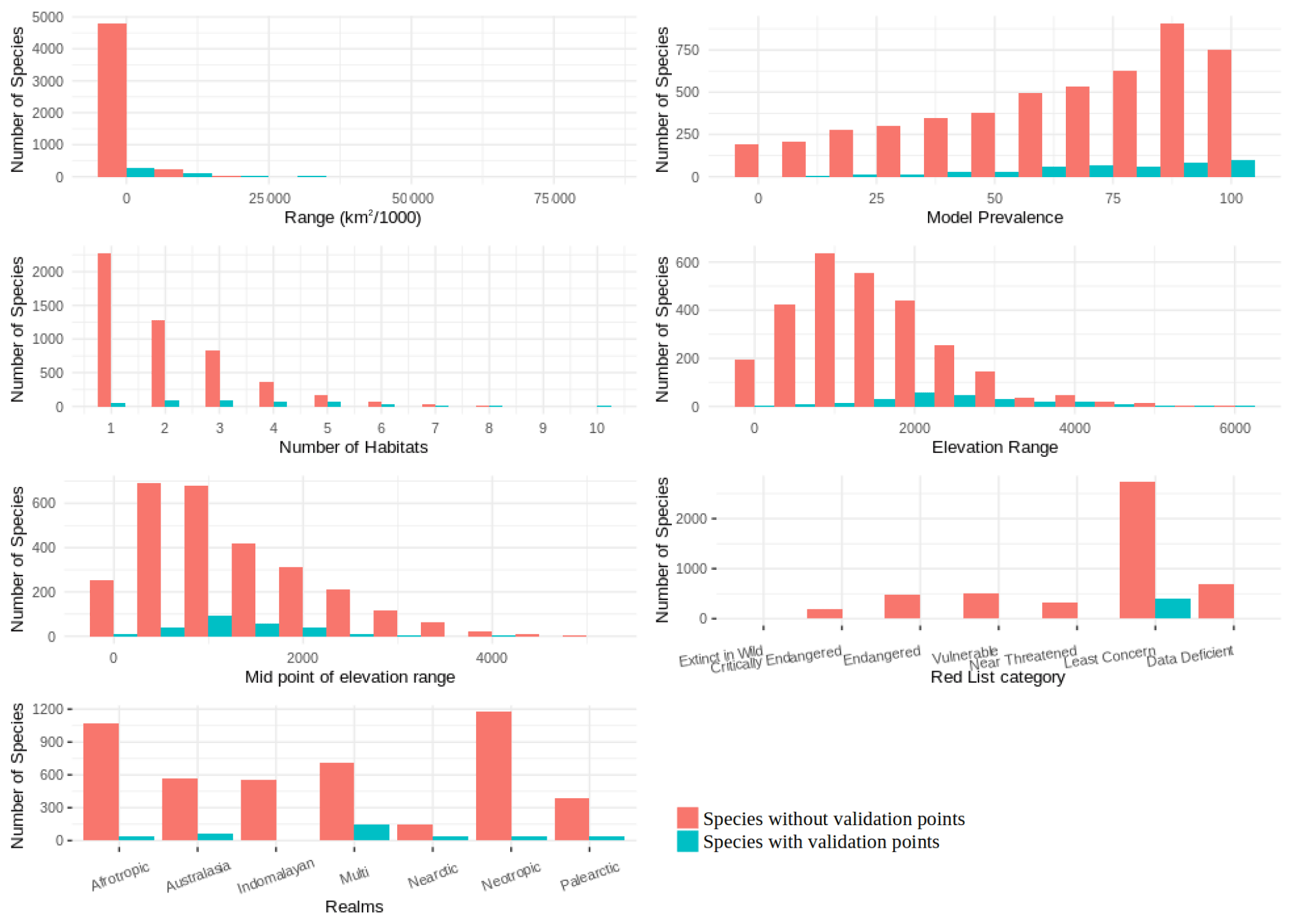
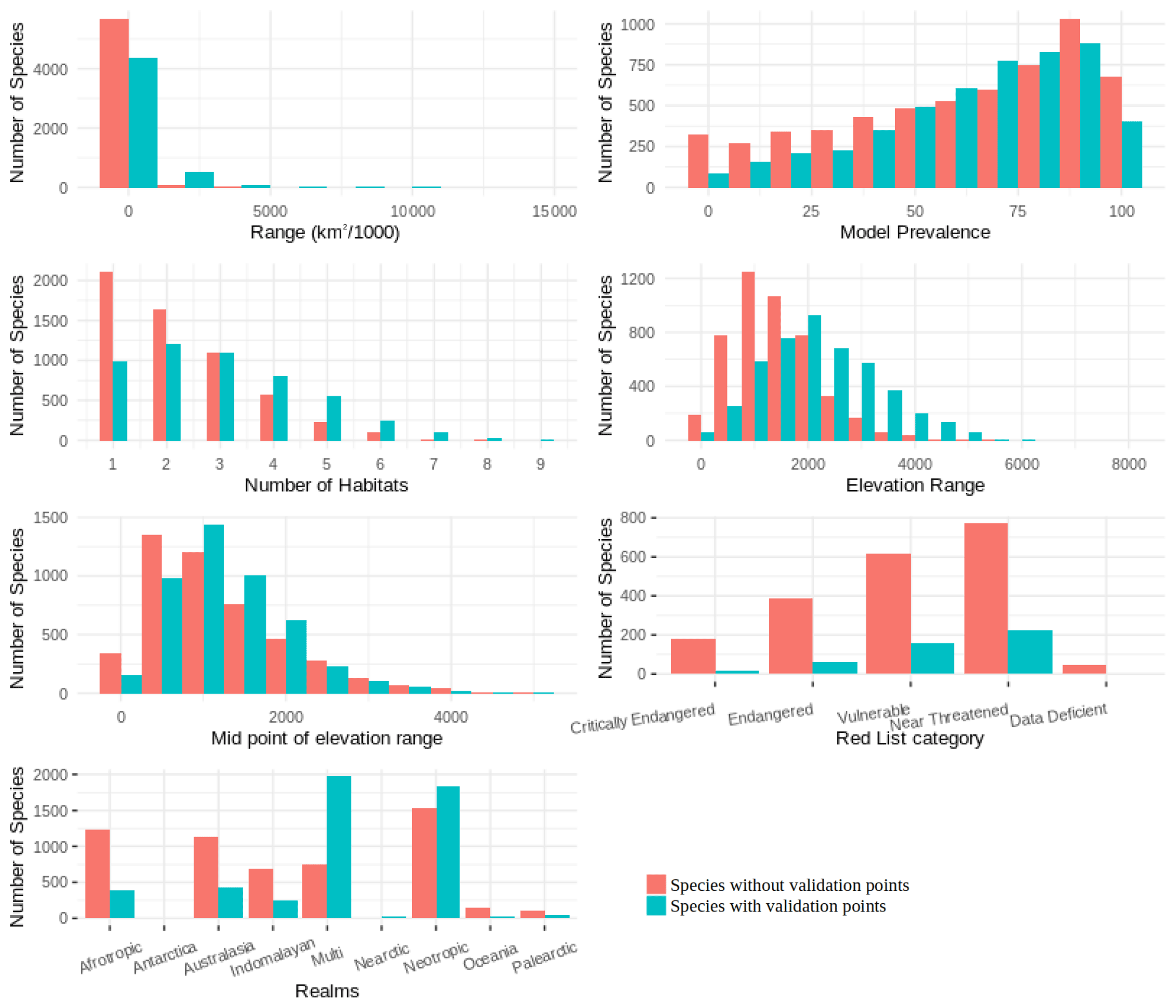
We found that for birds over 60 % of all families, genera and orders were represented in the sample included in the point validation and species from all biomes were represented but representation for mammals was lower, as expected due to the much lower proportion of mammal species for which point locality data were available (Fig. 5).
The validation points were spread across all of the variables and the majority of the subclasses (Figs. A6, A7). Species with validation points tended to have larger range sizes, wider elevation ranges and to be coded to more habitat classes than those without. Furthermore, validation points were not available for any critically endangered or endangered mammals as these species are rare in the wild.
3.3 Discussion
On comparing our point validation results with previous validation analysis of AOH maps, we found that the validation results are similar to or better than previous exercises. For mammals, Rondinini et al. (2011) evaluated AOH maps for 263 species at 300 m resolution, of which 241 (91.6 %) were better than random as compared to 95.0 % in our analysis. However, it should be noted that the mean model prevalence for the AOH maps of Rondinini et al. (2011) was 54.8 ± 21.5 SD as compared to 65.16 ± 25.42 for our AOH maps. The ratio of mean point prevalence to mean model prevalence for Rondinini et al. (2011) was 1.4 compared to 1.38 in our case. Ficetola et al. (2015) found that AOH for 94 % of 115 amphibian species used in the validation analysis were better than random with the mean model prevalence for species with validation points being 0.79 ± 0.21 SD. The ratio of mean point prevalence to mean model prevalence was 1.18 in this case.
Moreover, Catullo et al. (2008) found that 140 AOH maps out of 190 (73.7 %) South Asian mammal species gave positive validation results while Rondinini et al. (2005) found the mean proportion of suitable habitats correctly mapped inside the range for 181 species of African vertebrates was 0.55 ± 0.01 SE using presence-absence data sets. The high validation success in our analyses could be attributed to the use of a novel habitat-land cover model (Lumbierres et al., 2021a), the use of logistic regression models to identify systematic errors and the larger validation sample as compared with previous exercises. Furthermore, the underlying land cover map used in Lumbierres et al. (2021b), has the highest resolution among the global land cover maps providing more detailed land cover classification.
The point validation identified a small proportion of AOH maps which were no better than random. Some of these had high model prevalence. In such cases, point prevalence must be exceptionally high for the models to be better than random since even if the majority of point localities fall within the AOH these maps may perform no better than random. For the AOH maps which were no better than random and had low point prevalence, this was usually due to an apparent error in the coding of elevation range of the species, the areas inside the range of the species where the point localities fell being clipped out by what was assumed to be an erroneous elevation range. A list of species with probably erroneous elevation coding will be forwarded to IUCN Red List team for future corrections.
The AOH maps aim to minimize the commission errors known to be present in species ranges without increasing omission errors (Rondinini and Boitani, 2006). One of the limitations of this validation analysis is the inability to quantify the commission errors of the AOH maps as we do not have the true absence data of the species. Therefore, some uncertainty remains in AOH maps regarding the commission errors.
Also, there are some intrinsic errors in the models as identified by the logistic regression analysis. The species which are coded only to habitats like “shrubland” might have underrepresented model prevalence as discussed above. However, the number of AOH maps identified as lower outliers by the application of the logistic model was low for birds () and for mammals (), indicating that for the majority of AOH maps the observed model prevalence was fairly close to that predicted by the model.
The AOH maps validated in this paper is the largest validation done to date in terms of the number of species validated for birds and mammals. These maps will be freely available after the publication of Lumbierres et al. (2021b). We have also provided the metadata for all the species along with validation statistics in this paper, which can be used as a guideline by the users while using the AOH maps.
Figure A1AOH map for species Zimmerius chicomendesi. The species is coded against “forest” and “shrubland” habitats and the elevation range falls inside the IUCN range. However, the land cover inside this range map includes a high proportion of “herbaceous cover” land cover type which is not associated with “shrubland” habitat in the habitat-land cover association table. Therefore, the model prevalence of this AOH is much lower than expected.
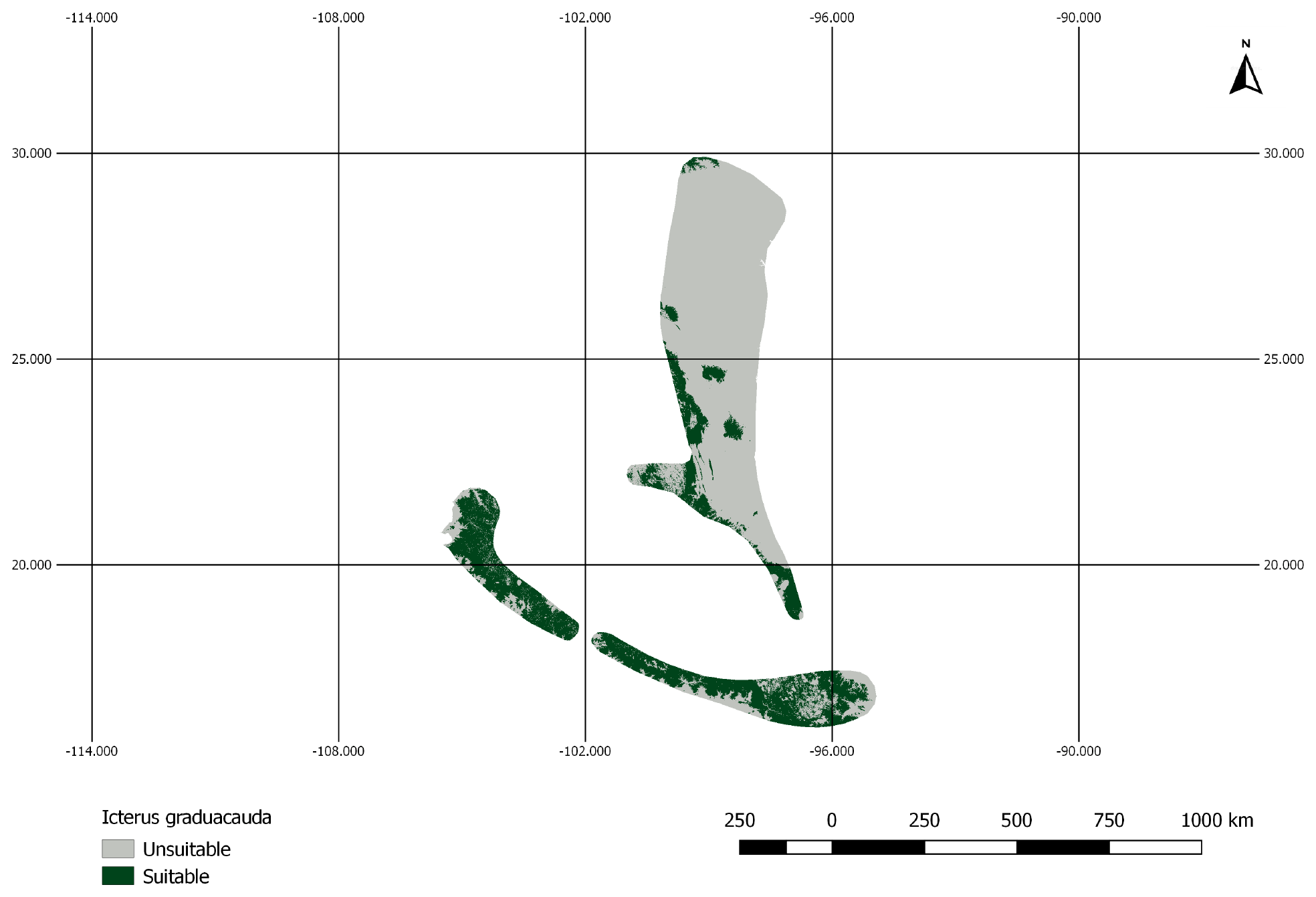
Figure A2AOH map for the species Icterus graduacauda. The IUCN range of the species does not cover much of the elevation range. Therefore, the model prevalence of this species is lower than estimated.

Figure A3AOH for the species Semnopithecus entellus. There is a large proportion of land cover class “cropland” inside the range map of this species. However, this species is not coded to habitats that are associated with the land cover “cropland”. Therefore, the model prevalence is lower than estimated.
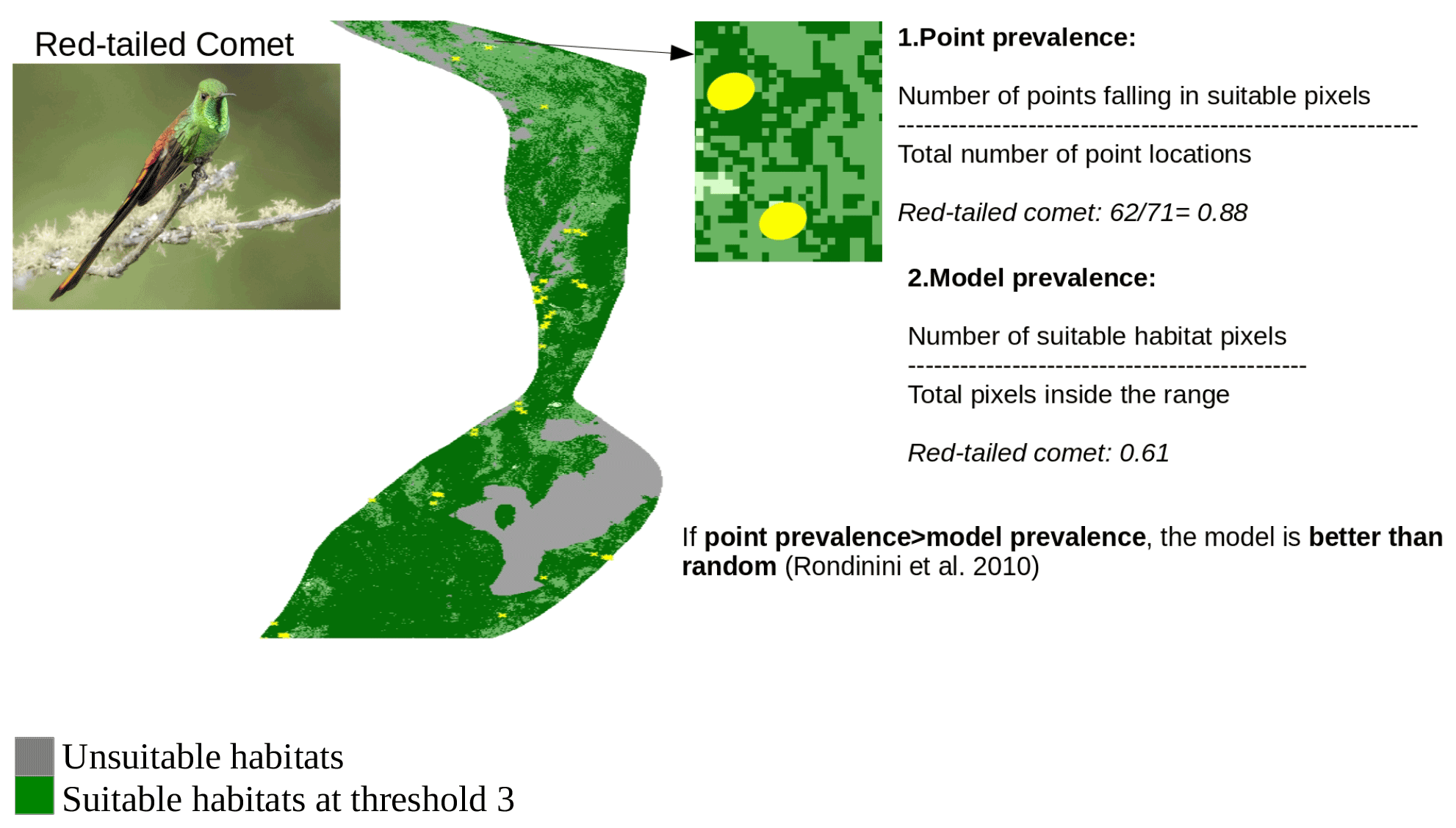
Figure A4Point validation of the AOH map of the species red-tailed comet using model and point prevalence. The yellow circles represent the buffered point localities of red-tailed comet. Image credit: Andres Vasquez Noboa, Macaulay Library ML 239910751.
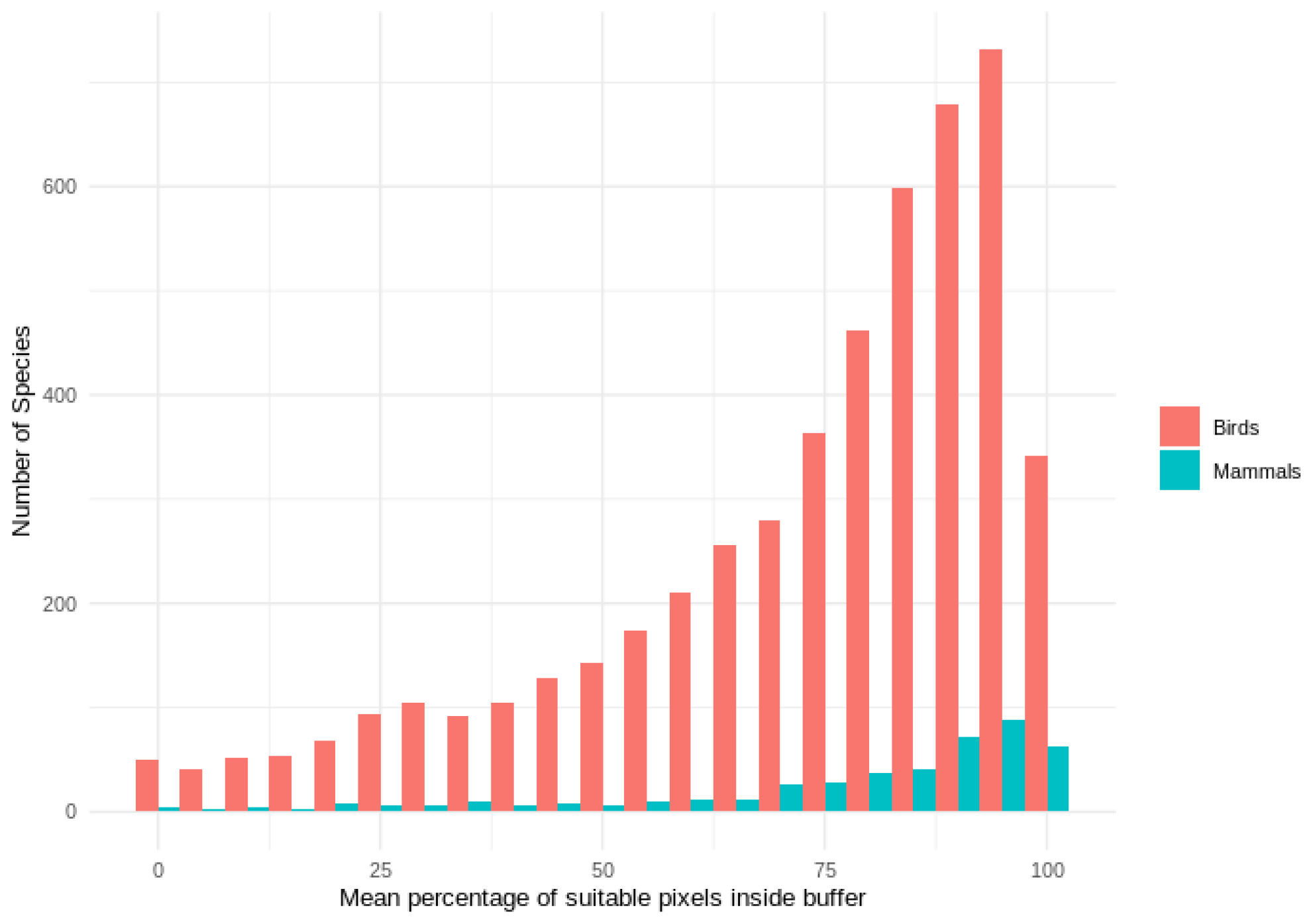
Figure A5Histogram of mean percentage of suitable AOH pixels inside the 300 m buffer for mammals and birds species used in point validation.
The point localities used in the validation analyses along with the metadata tables summarizing the validation analyses can be found at https://doi.org/10.5281/zenodo.5109073 (Dahal et al., 2021). The same DOI can be used to access the code used for validation and to also access some sample AOH maps which were validated.
PRD, PFD and CR conceptualized the idea. PRD and ML curated and did the formal data analysis. PRD led the manuscript writing with contributions from all the authors. PFD, CR, SHMB supervised the whole process.
The contact author has declared that neither they nor their co-authors have any competing interests.
Publisher’s note: Copernicus Publications remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
This research is part of the Inspire4Nature Innovative Training Network, funded by the European Union's Horizon 2020 research and innovation program under the Marie Skłodowska-Curie Actions program (grant no. 766417).
This paper was edited by Christian Folberth and reviewed by Everton Miranda and one anonymous referee.
Allouche, O., Tsoar, A., and Kadmon, R.: Assessing the accuracy of species distribution models: prevalence, kappa and the true skill statistic (TSS), J. Appl. Ecol., 43, 6, 1223–1232, https://doi.org/10.1111/j.1365-2664.2006.01214.x, 2006.
Bates, D., Mächler, M., Bolker, B., and Walker, S.: Fitting Linear Mixed-Effects Models Using lme4, J. Stat. Softw., 67, 1–48, https://doi.org/10.18637/jss.v067.i01, 2015.
BirdLife International and Handbook of the Birds of the World: Handbook of the Birds of the World and BirdLife International digital checklist of the birds of the world, Version 5, BirdLife International [data set], http://datazone.birdlife.org/userfiles/file/Species/Taxonomy/HBWBirdLife_Checklist_v5_Dec20.zip (last access: 3 March 2020), 2020.
Boyce, M. S., Vernier, P. R., Nielsen, S. E., and Schmiegelow, F. K.: Evaluating resource selection functions, Ecol. Model., 157, 281–300, https://doi.org/10.1016/S0304-3800(02)00200-4, 2002.
Brooks, T. M., Pimm, S. L., Akçakaya, H. R., Buchanan, G. M., Butchart, S. H., Foden, W., Hilton-Taylor, C., Hoffmann, M., Jenkins, C. N., Joppa, L., Li, B. V., Menon, V., Ocampo-Peñuela, N., and Rondinini, C.: Measuring Terrestrial Area of Habitat (AOH) and Its Utility for the IUCN Red List, Trends Ecol. Evol., 34, 977–986, https://doi.org/10.1016/j.tree.2019.06.009, 2019.
Buchhorn, M., Smets, B., Bertels, L., Lesiv, M., Tsendbazar, N., Herold, M., and Fritz, S.: Copernicus Global Land Service: Land Cover 100 m: epoch 2015: Globe, Dataset of the global component of the Copernicus Land Monitoring Service, Zenodo [data set], https://doi.org/10.5281/zenodo.3243509, 2019.
Catullo, G., Masi, M., Falcucci, A., Maiorano, L., Rondinini, C., and Boitani, L.: A gap analysis of Southeast Asian mammals based on habitat suitability models, Biol. Conserv., 141, 2730–2744, https://doi.org/10.1016/j.biocon.2008.08.019, 2008.
Chamberlain, S., Barve, V., Mcglinn, D., Oldoni, D., Desmet, P., Geffert, L., and Ram, K.: rgbif: Interface to the Global Biodiversity Information Facility API, R package version 3.5.2, Cran [software], https://CRAN.R-project.org/package=rgbif (last access: 1 January 2020), 2021.
Cornell Lab of Ornithology.: eBird Basic Dataset, Version: EBD_Jan 2020, eBird [data set], http://www.ebird.com/data/download (last access: 14 June 2021), 2020.
Dahal, P. R., Lumbierres, M., Butchart, S. H. M., Donald, P. F., and Rondinini, C.: Data used, summary and codes: A validation standard for Area of Habitat maps for terrestrial birds and mammals, Zenodo [code and data set], https://doi.org/10.5281/zenodo.5109073, 2021.
Ficetola, G. F., Rondinini, C., Bonardi, A., Baisero, D., and Padoa Schioppa, E.: Habitat availability for amphibians and extinction threat: A global analysis, Divers. Distrib., 21, 302–311, https://doi.org/10.1111/ddi.12296, 2015.
GBIF.org.: Derived dataset GBIF.org Filtered export of GBIF occurrence data, GBIF.org [data set], https://doi.org/10.15468/dd.mezbz7, 2021.
GRASS Development Team: Geographic Resources Analysis Support System (GRASS) Software, Version 7.2. Open Source Geospatial Foundation, Electronic document, http://grass.osgeo.org (last access: 2 June 2020), 2017.
IUCN: Habitats Classification Scheme (Version 3.1), IUCN, Gland, 2012.
IUCN: The IUCN Red List of Threatened Species, Version 2020-2, IUCN, Gland, 2020.
Jenkins, C. N. and Giri, C.: Protection of mammal diversity in Central America, Conserv. Biol., 22, 1037–1044, https://doi.org/10.1111/j.1523-1739.2008.00974.x, 2008.
Jung, M., Dahal, P. R., Butchart, S. H. M., Donald, P. F., De Lamo, X., Lesiv, M., Kapos, V., Rondinini, C., and Visconti, P.: A global map of terrestrial habitat types, Scientific Data, 7, 256, https://doi.org/10.1038/s41597-020-00599-8, 2020.
Lüdecke, D., Waggoner, P. D., and Makowski, D.: insight: A Unified Interface to Access Information from Model Objects in R, Journal of Open Source Software, 4, 1412, https://doi.org/10.21105/joss.01412, 2019.
Lumbierres, M., Dahal, P. R., Di Marco, M., Butchart, S. H., Donald, P. F., and Rondinini, C.: Translating habitat class to land cover to map area of habitat of terrestrial vertebrates, Conserv. Biol., 36, e13851, https://doi.org/10.1111/cobi.13851, 2021a.
Lumbierres, M., Dahal, P. R., Di Marco, M., Butchart, S. H., Donald, P. F., and Rondinini, C.: Area of Habitat maps for the world’s terrestrial birds and mammals, bioRxiv [preprint], https://doi.org/10.1101/2022.05.13.489640, 2021b.
R Core Team.: R: A language and environment for statistical computing, R Foundation for Statistical Computing, https://www.R-project.org/ (last access: 1 January 2020), 2018.
Rondinini, C. and Boitani, L.: Differences in the umbrella effects of African amphibians and mammals based on two estimators of the area of occupancy, Conserv. Biol., 20, 170–179, https://doi.org/10.1111/j.1523-1739.2005.00299.x, 2006.
Rondinini, C., Stuart, S., and Boitani, L..: Habitat suitability models and the shortfall in conservation planning for African vertebrates, Conserv. Biol., 19, 1488–1497, https://doi.org/10.1111/j.1523-1739.2005.00204.x, 2005.
Rondinini, C., Di Marco, M., Chiozza, F., Santulli, G., Baisero, D., Visconti, P., Hoffmann, M., Schipper, J., Stuart, S. N., Tognelli, M. F., Amori, G., Falcucci, A., Maiorano, L., and Boitani, L.: Global habitat suitability models of terrestrial mammals, Philos. T. R. Soc. B., 366, 2633–2641, https://doi.org/10.1098/rstb.2011.0113, 2011.
Stoms, D. M., Davis, F. W., and Cogan, C. B.: Sensitivity of wildlife habitat models to uncertainties in GIS data, Photogramm. Eng. Rem. S., 58, 843–850, 1992.
Strimas-Mackey, M., Miller, E., and Hochachka, W.: auk: eBird Data Extraction and Processing with AWK, R package version 0.3.0, https://cornelllabofornithology.github.io/auk/ (last access: 1 May 2020), 2018.
Sullivan, L. B., Wood, C. L., Iliff, M. J., Bonney, R. E., Fink, D., and Kelling, S.: eBird: A citizen-based bird observation network in the biological sciences, Biol. Conserv., 142, 2282–2292, https://doi.org/10.1016/j.biocon.2009.05.006, 2009.
Tracewski, L., Butchart, S. H., Di Marco, M., Ficetola, G. F., Rondinini, C., Symes, A., Wheatley, H., Beresford, A. E., and Buchanan, G. M.: Toward quantification of the impact of 21st-century deforestation on the extinction risk of terrestrial vertebrates, Conserv. Biol., 30, 1070–1079, https://doi.org/10.1111/cobi.12715, 2016.
Wilcox, R. R.: Introduction to robust estimation and hypothesis testing: 4th edn., Elsevier, 713 Waltham, Massachusetts, USA, https://doi.org/10.1016/C2010-0-67044-1, 2017.